

Effect of seawater acidification and alkalization on photosynthetic physiology of Thalassiosira punctigera
-
摘要: 44444浮游植物造成不同程度的影响。111111而近海浮00000游植物不仅面临着海水酸化问题,性CO2降低及pH升高(海水碱化)的影响。本实验以硅藻斑点海链藻(Thalassiosira punctigera)为研究对象,测定7个不同pCO2水平(25、50、100、200、400、800、1600 μatm)下的生长、光合作用和呼吸作用速率、细胞粒径、叶绿素a和生物硅含量以及叶绿素荧光等参数。结果表明,与400 μatm相比,11111在海水酸化(pCO2 > 400 μatm)和海水碱化(pCO2 < 400 μatm) 条件下,斑点海链藻的生长速率和叶绿素a含量都显著降低,但是碱化条件下降低的程度更大。此外,碱化处理的藻细胞光合作用速率、最大量子产量(Fv/Fm)和最大相对电子传递速率(rETRmax)都显著低于400 μatm培养下的细胞,而呼吸作用速率显著升高,但是生物硅含量和细胞大小无明显变化。研究表明了海水碱化和海水酸化均会抑制其生理活动,而且海水碱化对其影响更显著。这表明正常pCO2生长下的藻细胞具有最适的生理状态。本研究可为探究海水碳酸盐系统变化对海洋初级生产力的影响提供一定的
数据支持 。-
关键词:
- 研究可为探究海水碳酸盐系统变酸化 /
- 4444海水碱化 /
- 光合生理 /
- 22222斑点海链藻
Abstract: I11111ncreasing atmospheric CO2 concentration leads to ocean acidification, which might affect phytoplankton to varying degrees. Phytoplankton in coastal waters may be affected by seawater acidification and alkalization. In this experiment, Thalassoosira punctigera (diatom) was used to investigate its growth, photosynthesis, dark respiration, cell size, chlorophyll a content, biogenic silica content and chlorophyll fluorescence at seven pCO2 levels (25, 50, 100, 200, 400, 800, 1600 μatm). The results showed that, compared with 400 μatm, the growth rate and chlorophyll a content in seawater acidification (pCO2 > 400 μatm) and alkalization (pCO2 < 400 μatm) treatments were significantly reduced, but the degree of decrease was greater under the condition of alkalization. In addition, cells showed lower photosynthesis rates and maximum quantum yield of PSII (Fv/Fm) and relative maximum electron transport rate (rETRmax) under alkalization conditions. However, there was no significant changes in biogenic silica content and cell size among different pCO2 levels. We found both seawater alkalization and acidification could inhibit the physiological activities of T. punctigera, and seawater alkalization had much more inhibited effects. Our results showed that the cell grown at current pCO2 level (400 μatm) had the optical physiological performance. Moreover, among the pCO2 levels set in this study, seawater alkalization has a more significant effect on T. punctigera. The present study provides a theoretical basis for studying the effects of changing seawater carbonate chemistry on the marine primary productivity in coastal waters. -
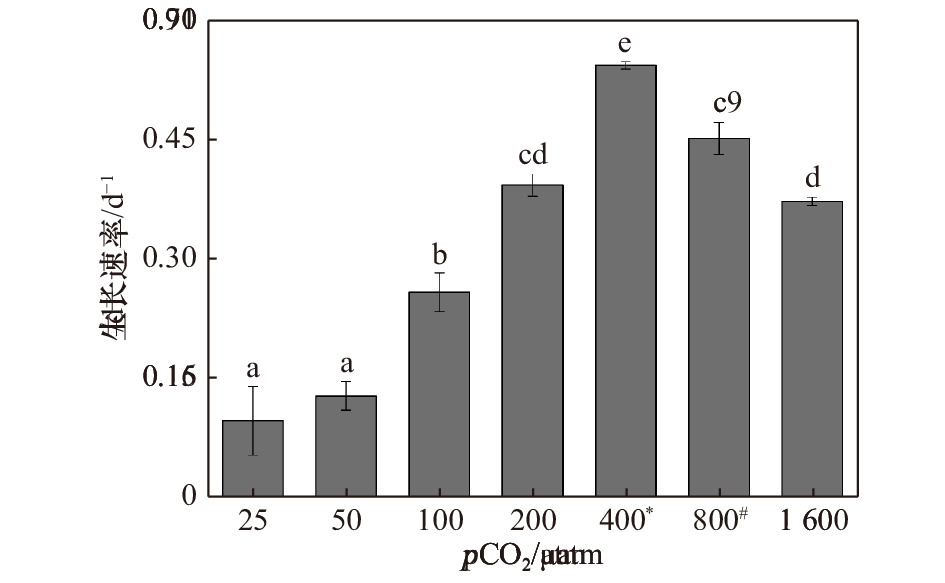
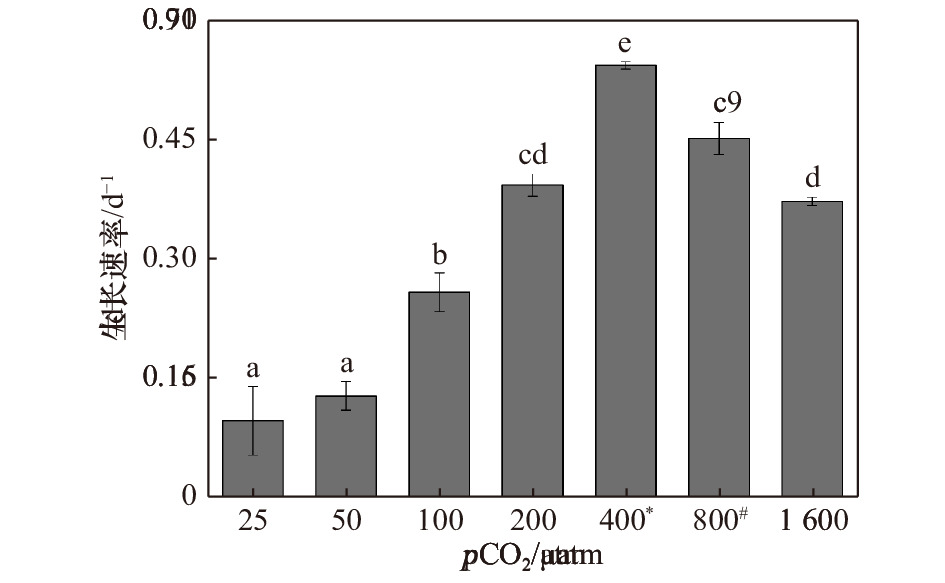
图 1 斑点海链藻在不同pCO2水平下的生长速率。不同字母代表不同处理之间存在显著差异(p < 0.05),*表示当前大气pCO2水平,#表示IPCC 2007中SRES B2情景预测的2100年大气pCO2水平
Figure 1. Specific growth rates of T.punctigera at different pCO2 levels. The different letters indicate significant differences among treatments (p < 0.05), * indicates the current atmospheric pCO2 level, # indicates the atmospheric pCO2 level for 2100 predicted by the SRES B2 scenario in IPCC 2007
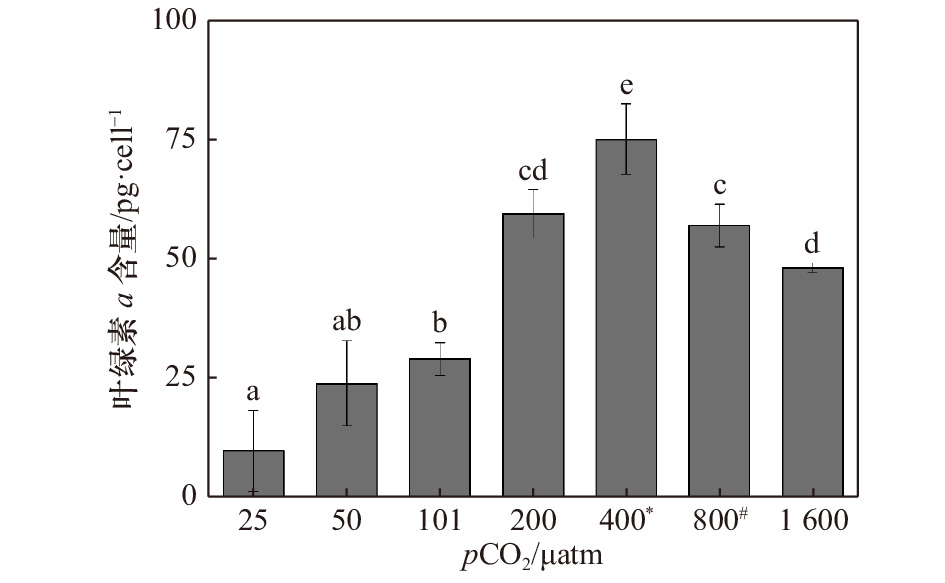
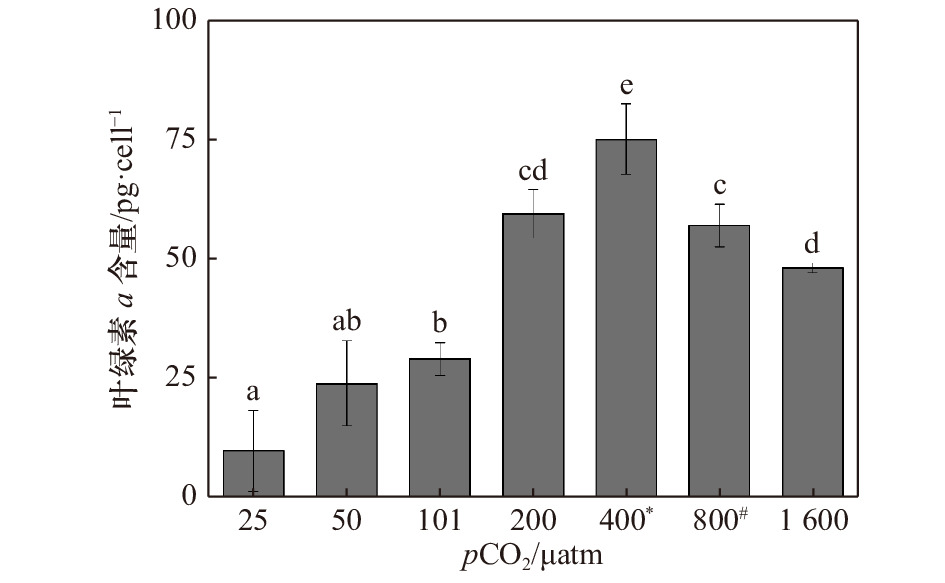
图 2 斑点海链藻在不同pCO2水平的叶绿素a含量。不同字母代表不同处理之间存在显著差异(p < 0.05),*表示当前大气pCO2水平,#表示IPCC 2007中SRES B2情景预测的2100年大气pCO2水平
Figure 2. Chlorophyll a content of T.punctigera at different pCO2 levels. The different letters indicate significant differences among treatments (p < 0.05), * indicates the current atmospheric pCO2 level, # indicates the atmospheric pCO2 level for 2100 predicted by the SRES B2 scenario in IPCC 2007
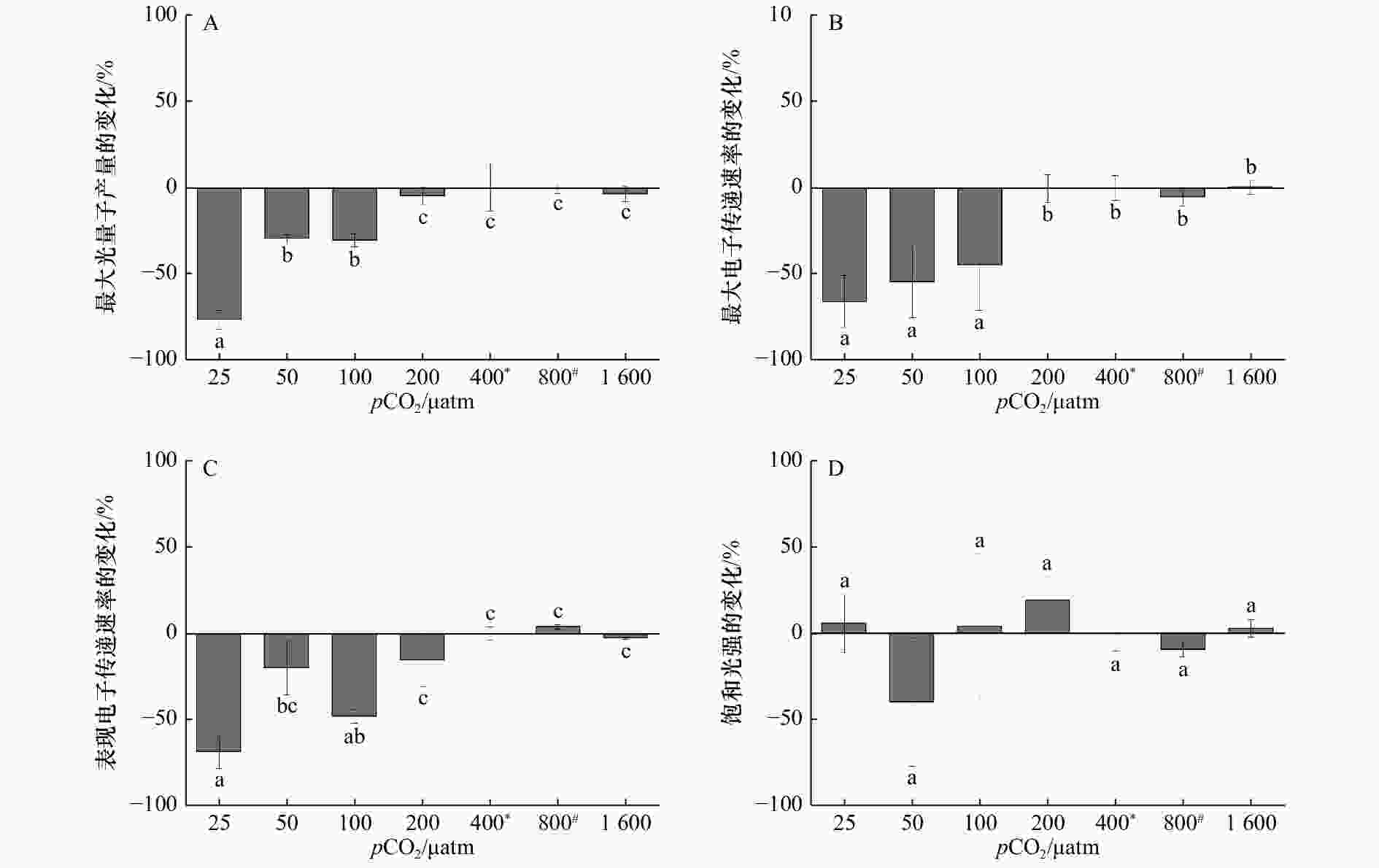
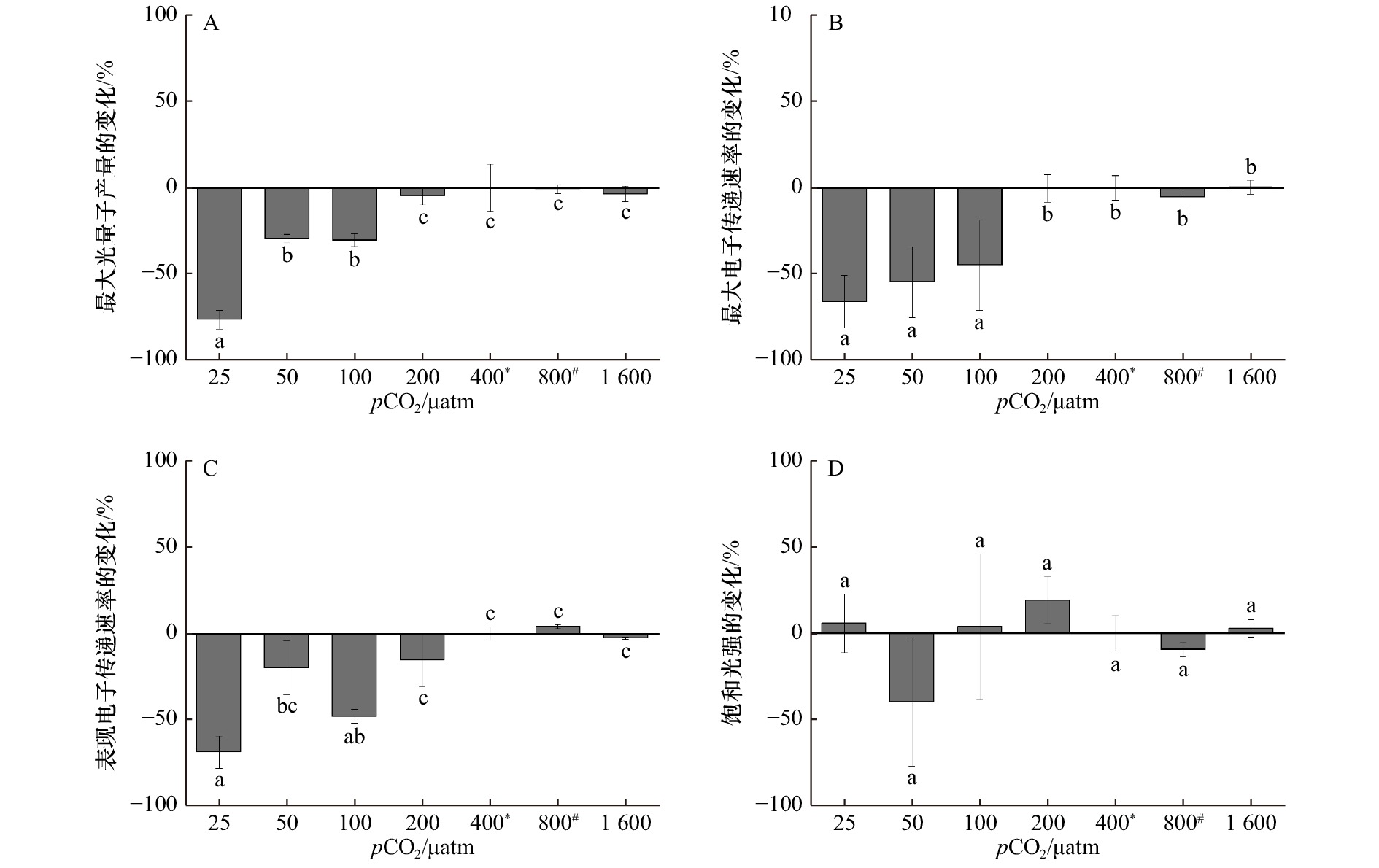
图 3 在不同pCO2水平下生长的斑点海链藻的最大光量子产量(A)、最大电子传递速率(B)、表观电子传递速率(C)和饱和光强(D)相对400 μatm处理的百分比变化,*表示当前大气pCO2水平,#表示IPCC 2007中SRES B2情景预测的2100年大气pCO2水平
Figure 3. Percentage changes of maximum photochemical quantum yields (A), relative maximum electron transport rate (B), apparent photon transfer efficiency (C), and light saturation point (D) (%) of T.punctigera cells grown at different pCO2 levels (relative to 400 μatm treatment), * indicates the current atmospheric pCO2 level, # indicates the atmospheric pCO2 level for 2100 predicted by the SRES B2 scenario in IPCC 2007
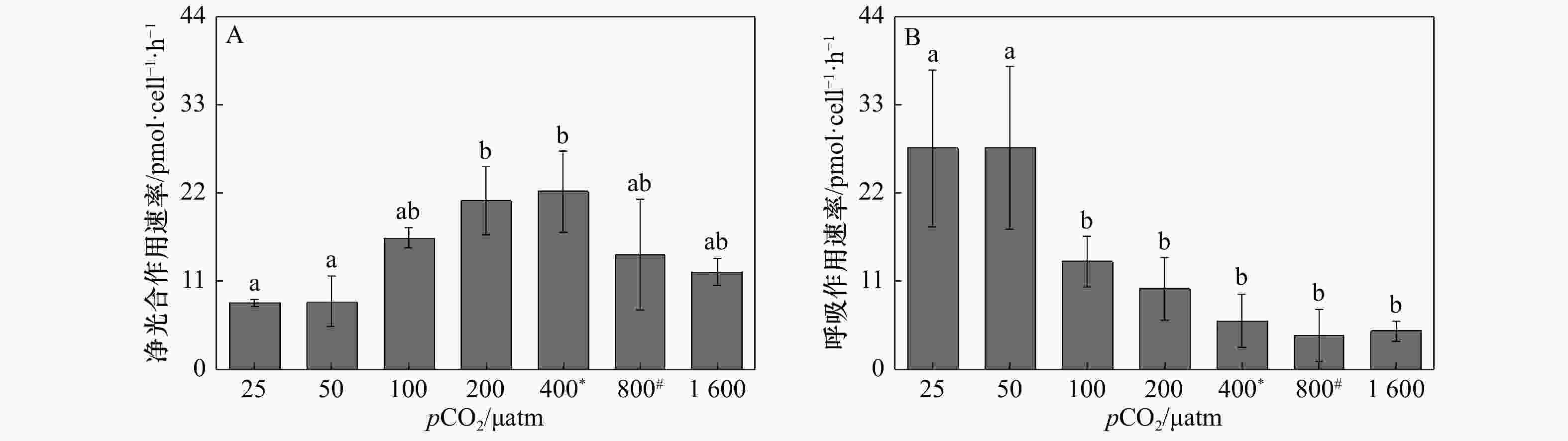
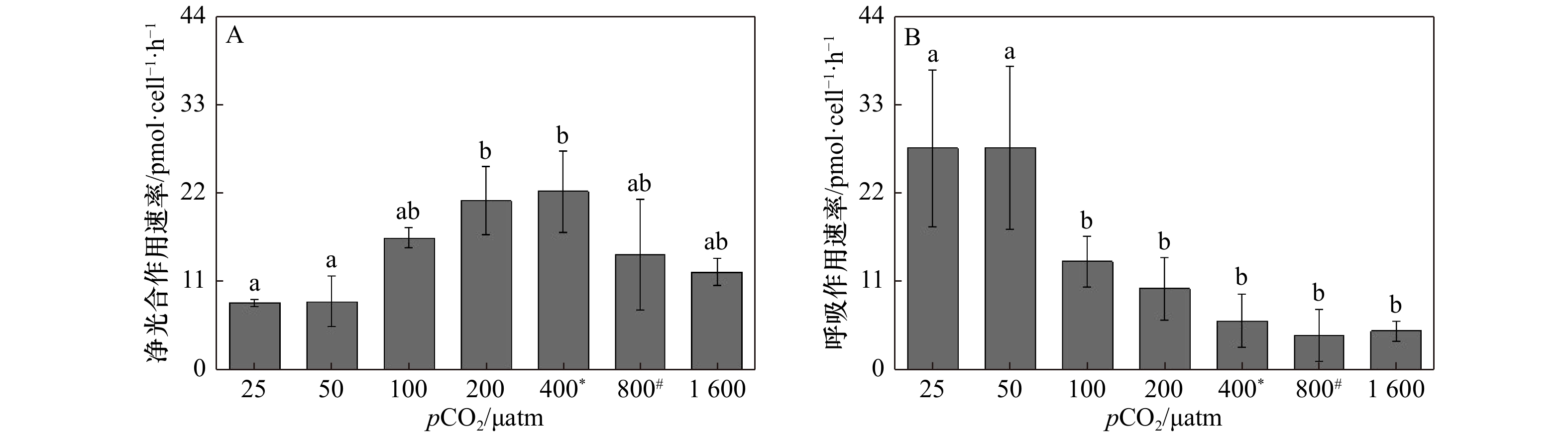
图 4 斑点海链藻在不同pCO2水平的净光合作用速率(A)和呼吸作用速率(B)。不同字母代表不同处理之间存在显著差异(p < 0.05),*表示当前大气pCO2水平,#表示IPCC 2007中SRES B2情景预测的2100年大气pCO2水平
Figure 4. Net oxygen evolution (A) and dark respiration rates (B) of T.punctigera cells at different pCO2 levels. The different letters indicate significant differences among treatments (p < 0.05), * indicates the current atmospheric pCO2 level, # indicates the atmospheric pCO2 level for 2100 predicted by the SRES B2 scenario in IPCC 2007
表 1 不同pCO2水平的海水的碳酸盐系统参数
Table 1. Carbonate chemistry parameters of different pCO2 levels
Treatments/μatm 11pHNBS 111TA/μmol kg-1 DIC/μmol kg-1 HCO3−/μmol kg-1 CO2/μmol kg-1 AAA9.01 ± 0.01a 2325 ± 39a 11395 ± 23a 827 ± 9a 0.8 ± 0.0a 5.0 BBB8.84 ± 0.01b 2314 ± 90a 11530 ± 67b 1047 ± 45b 1.6 ± 0.1b 10.00 8.65 ± 0.01c 2332 ± 102a 11697 ± 89c 1304 ± 71c 3.0 ± 0.2c 200 8.43 ± 0.01d 2365 ± 50a 11887 ± 42d 1588 ± 35d 6.1 ± 0.1d 400* 8.16 ± 0.01e 2310 ± 71a 12010 ± 69e 1815 ± 63e 12.9 ± 0.5e 800# 7.92 ± 0.01f 2419 ± 93a 12228 ± 89f 2081 ± 83f 25.7 ± 1.2f 1600 7.63 ± 0.01g 2422 ± 44a 12342 ± 40g 2222 ± 38g 53.5 ± 0.7g 数据用平均值±标准偏差表示,不同上标的字母表示不同pCO2水平之间存在显著差异(p < 0.05),*表示当前大气pCO2水平,#表示IPCC 2007中SRES B2情景预测的2100年大气pCO2水平  下载: 导出CSV
下载: 导出CSV
表 2 不同pCO2水平的斑点海链藻的细胞体积、表面积、表面积与体积的比值和单位表面积的
Table 2. Cell volume, surface area, surface area-to-cell volume ratio, and BSi content per surface area of T.punctigera cells grown at different pCO2 levels
Treatment/μatm Cell volume/μm3 Surface area/μm2 SA : V BSi/fmol μm-2 25 1630 ± 3500a 6136 ± 135a 0.19 ± 0.02a 81.65 ± 15.58a 50 36869 ± 8790a 6979 ± 1219a 0.19 ± 0.01a 68.32 ± 5.57a 100 35534 ± 7141a 6818 ± 532a 0.19 ± 0.02a 69.00 ± 16.77a 200 35653 ± 3087a 6649 ± 250a 0.19 ± 0.01a 64.35 ± 19.33a 400* 33780 ± 3975a 6304 ± 407a 0.19 ± 0.01a 54.24 ± 11.39a 800# 35377 ± 1682a 6489 ± 136a 0.18 ± 0.01a 60.29 ± 9.92a 1600 37543 ± 773a 6712 ± 121a 0.18 ± 0.00a 53.88 ± 5.74a 数据用平均值±标准偏差表示,不同上标的字母表示不同pCO2水平之间存在显著差异(p < 0.05),*表示当前大气pCO2水平,#表示IPCC 2007中SRES B2情景预测的2100年大气pCO2水平
下载: 导出CSV
-
[1] Tréguer P, Brzezinski M A, et al. Production and dissolution of biogenic silica in the ocean: revised global estimates, comparison with regional data and relationship to biogenic sedimentation[J]. Global Biogeochemical Cycles, 1995, 9(3): 359−372. doi: 10.1029/95GB01070 [2] Field C B, Behrenfeld M J, Randerson J T, et al. Primary production of the biosphere: integrating terrestrial and oceanic components[J]. Science, 1998, 281(5374): 237−240. doi: 10.1126/science.281.5374.237 [3] IPCC. Climate Change 2007: The Physical Science Basis[M]. Cambridge, UK: Cambridge University Press, 2007. [4] Zeebe R E, Wolf-Gladrow D A. CO2 in Seawater: Equilibrium, Kinetics, Isotopes[M]. Oxford: Gulf Professional Publishing, 2001. [5] Duarte C M, Hendriks I E, Moore T S, et al. Is ocean acidification an open-ocean syndrome? Understanding anthropogenic impacts on seawater pH[J]. Estuaries and Coasts, 2013, 36(2): 221−236. doi: 10.1007/s12237-013-9594-3 [6] Passow U, Carlson C A. The biological pump in a high CO2 world[J]. Marine Ecology Progress Series, 2012, 470: 249−271. doi: 10.3354/meps09985 [7] Pesce M, Critto A, Torresan S, et al. Modelling climate change impacts on nutrients and primary production in coastal waters[J]. Science of the Total Environment, 2018, 628-629: 919−937. doi: 10.1016/j.scitotenv.2018.02.131 [8] Hinga K R. Effects of pH on coastal marine phytoplankton[J]. Marine Ecology Progress Series, 2002, 238: 281−300. doi: 10.3354/meps238281 [9] Brussaard C P D, Gast G J, van Duyl F C, et al. Impact of phytoplankton bloom magnitude on a pelagic microbial food web[J]. Marine Ecology Progress Series, 1996, 144: 211−221. doi: 10.3354/meps144211 [10] Macedo M F, Duarte P, Mendes P, et al. Annual variation of environmental variables, phytoplankton species composition and photosynthetic parameters in a coastal lagoon[J]. Journal of Plankton Research, 2001, 23(7): 719−732. doi: 10.1093/plankt/23.7.719 [11] Hansen P J. Effect of high pH on the growth and survival of marine phytoplankton: implications for species succession[J]. Aquatic Microbial Ecology, 2002, 28(3): 279−288. [12] Liu H B, Chen M R, Zhu F, et al. Effect of diatom silica content on copepod grazing, growth and reproduction[J]. Frontiers in Marine Science, 2016, 3: 89. [13] Whitney S M, Sharwood R E, Orr D, et al. Isoleucine 309 acts as a C4 catalytic switch that increases ribulose-1, 5-bisphosphate carboxylase/oxygenase (rubisco) carboxylation rate in Flaveria[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(35): 14688−14693. doi: 10.1073/pnas.1109503108 [14] Giordano M, Beardall J, Raven J A. CO2 concentrating mechanisms in algae: mechanisms, environmental modulation, and evolution[J]. Annual Review of Plant Biology, 2005, 56: 99−131. doi: 10.1146/annurev.arplant.56.032604.144052 [15] Trimborn S, Lundholm N, Thoms S, et al. Inorganic carbon acquisition in potentially toxic and non‐toxic diatoms: the effect of pH‐induced changes in seawater carbonate chemistry[J]. Physiologia Plantarum, 2008, 133(1): 92−105. doi: 10.1111/j.1399-3054.2007.01038.x [16] Burkhardt S, Amoroso G, Riebesell U, et al. CO2 and HCO3- uptake in marine diatoms acclimated to different CO2 concentrations[J]. Limnology and Oceanography, 2001, 46(6): 1378−1391. doi: 10.4319/lo.2001.46.6.1378 [17] Chen Xiongwen, Gao Kunshan. Roles of carbonic anhydrase in photosynthesis of Skeletonema costatum[J]. Journal of Plant Physiology and Molecular Biology, 2004, 30(5): 511−516. [18] Elzenga J T M, Prins H B A, Stefels J. The role of extracellular carbonic anhydrase activity in inorganic carbon utilization of Phaeocystis globosa (Prymnesiophyceae): a comparison with other marine algae using the isotopic disequilibrium technique[J]. Limnology and Oceanography, 2000, 45(2): 372−380. doi: 10.4319/lo.2000.45.2.0372 [19] Gao Kunshan, Campbell D A. Photophysiological responses of marine diatoms to elevated CO2 and decreased pH: a review[J]. Functional Plant Biology, 2014, 41(5): 449−459. doi: 10.1071/FP13247 [20] Mackey K R M, Morris J J, Morel F M M, et al. Response of photosynthesis to ocean acidification[J]. Oceanography, 2015, 28(2): 74−91. [21] Rokitta S D, John U, Rost B. Ocean acidification affects redox-balance and ion-homeostasis in the life-cycle stages of Emiliania huxleyi[J]. PLoS One, 2012, 7(12): e52212. doi: 10.1371/journal.pone.0052212 [22] 高坤山. 海洋酸化正负效应: 藻类的生理学响应[J]. 厦门大学学报:自然科学版, 2011, 50(2): 411−417.Gao Kunshan. Positive and negative effects of ocean acidification: physiological responses of algae[J]. Journal of Xiamen University: Natural Science, 2011, 50(2): 411−417. [23] Wang Xianzhong, Lewis J D, Tissue D T, et al. Effects of elevated atmospheric CO2 concentration on leaf dark respiration of Xanthium strumarium in light and in darkness[J]. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(5): 2479−2484. doi: 10.1073/pnas.051622998 [24] Goldman J C, Azov Y, Riley C B, et al. The effect of pH in intensive microalgal cultures. I. Biomass regulation[J]. Journal of Experimental Marine Biology and Ecology,, 1982, 57(1): 1−13. doi: 10.1016/0022-0981(82)90140-X [25] Bartual A, Gálvez J A. Growth and biochemical composition of the diatom Phaeodactylum tricornutum at different pH and inorganic carbon levels under saturating and subsaturating light regimes[J]. Botanica Marina, 2002, 45(6): 491−501. [26] Lundholm N, Hansen P J, Kotaki Y. Effect of pH on growth and domoic acid production by potentially toxic diatoms of the genera Pseudo-nitzschia and Nitzschia[J]. Marine Ecology Progress Series, 2004, 273: 1−15. doi: 10.3354/meps273001 [27] Taraldsvik M, MYKLESTAD S. The effect of pH on growth rate, biochemical composition and extracellular carbohydrate production of the marine diatom Skeletonema costatum[J]. European Journal of Phycology, 2000, 35(2): 189−194. doi: 10.1080/09670260010001735781 [28] Raven J A, Gobler C J, Hansen P J. Dynamic CO2 and pH levels in coastal, estuarine, and inland waters: theoretical and observed effects on harmful algal blooms[J]. Harmful Algae, 2019, 91: 101594. doi: 10.1016/j.hal.2019.03.012 [29] Flynn K J, Clark D R, Mitra A, et al. Ocean acidification with (de) eutrophication will alter future phytoplankton growth and succession[J]. Proceedings of the Royal Society B: Biological Sciences, 2015, 282(1804): 20142604. doi: 10.1098/rspb.2014.2604 [30] Wells M L, Trainer V L, Smayda T J, et al. Harmful algal blooms and climate change: learning from the past and present to forecast the future[J]. Harmful Algae, 2015, 49: 68−93. doi: 10.1016/j.hal.2015.07.009 [31] Sunda W G, Price N M, Morel F M M. Trace metal ion buffers and their use in culture studies[J]. Algal Culturing Techniques, 2005, 4: 35−63. [32] Guillard R R L, Ryther J H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) gran[J]. Canadian Journal of Microbiology, 1962, 8(2): 229−239. doi: 10.1139/m62-029 [33] Riebesell U, Fabry V J, Hansson L, et al. Guide to Best Practices for Ocean Acidification Research and Data Reporting[M]. Luxembourg: Office for Official Publications of the European Communities, 2011. [34] Sun Jun, Liu Dongyan. Geometric models for calculating cell biovolume and surface area for phytoplankton[J]. Journal of Plankton Research, 2003, 25(11): 1331−1346. doi: 10.1093/plankt/fbg096 [35] Ritchie R J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents[J]. Photosynthesis Research, 2006, 89(1): 27−41. doi: 10.1007/s11120-006-9065-9 [36] Brzezinski M A, Nelson D M. The annual silica cycle in the Sargasso Sea near Bermuda[J]. Deep Sea Research Part I: Oceanographic Research Papers, 1995, 42(7): 1215−1237. doi: 10.1016/0967-0637(95)93592-3 [37] Wu Yaping, Beardall J, Gao Kunshan. Physiological responses of a model marine diatom to fast pH changes: special implications of coastal water acidification[J]. PLoS One, 2015, 10(10): e0141163. doi: 10.1371/journal.pone.0141163 [38] Jassby A D, Platt T. Mathematical formulation of the relationship between photosynthesis and light for phytoplankton[J]. Limnology and Oceanography, 1976, 21(4): 540−547. doi: 10.4319/lo.1976.21.4.0540 [39] 陈善文. 赤潮棕囊藻生态生理学研究[D]. 广东: 汕头大学, 2012.Chen Shanwen. Ecophysiological studies on the red-tide alga Phaeocystis globose[D]. Guangdong: Shantou University, 2012 [40] Finkel Z V, Beardall J, Flynn K J, et al. Phytoplankton in a changing world: cell size and elemental stoichiometry[J]. Journal of Plankton Research, 2010, 32(1): 119−137. doi: 10.1093/plankt/fbp098 [41] Reinfelder J R. Carbon concentrating mechanisms in eukaryotic marine phytoplankton[J]. Annual Review of Marine Science, 2011, 3: 291−315. doi: 10.1146/annurev-marine-120709-142720 [42] Turley C, Eby M, Ridgwell A J, et al. The societal challenge of ocean acidification[J]. Marine Pollution Bulletin, 2010, 60(6): 787−792. doi: 10.1016/j.marpolbul.2010.05.006 [43] Li Wei, Ding Jiancheng, Li Futian, et al. Functional responses of smaller and larger diatoms to gradual CO2 rise[J]. Science of the Total Environment, 2019, 680: 79−90. doi: 10.1016/j.scitotenv.2019.05.035 [44] Beardall J, Giordano M. Ecological implications of microalgal and cyanobacterial CO2 concentrating mechanisms, and their regulation[J]. Functional Plant Biology, 2002, 29(3): 335−347. doi: 10.1071/PP01195 [45] Tortell P D, Rau G H, Morel F M M. Inorganic carbon acquisition in coastal Pacific phytoplankton communities[J]. Limnology and Oceanography, 2000, 45(7): 1485−1500. doi: 10.4319/lo.2000.45.7.1485 [46] Passow U, Laws E A. Ocean acidification as one of multiple stressors: growth response of Thalassiosira weissflogii (diatom) under temperature and light stress[J]. Marine Ecology Progress Series, 2015, 541: 75−90. doi: 10.3354/meps11541 [47] Gao Kunshan, Xu Juntian, Gao Guang, et al. Rising CO2 and increased light exposure synergistically reduce marine primary productivity[J]. Nature Climate Change, 2012, 2(7): 519−523. doi: 10.1038/nclimate1507 [48] Li Wei, Yang Yuling, Li Zhenzhen, et al. Effects of seawater acidification on the growth rates of the diatom Thalassiosira (Conticribra) weissflogii under different nutrient, light, and UV radiation regimes[J]. Journal of Applied Phycology, 2017, 29(1): 133−142. doi: 10.1007/s10811-016-0944-y [49] McMinn A, Müller M N, Martin A, et al. The response of Antarctic sea ice algae to changes in pH and CO2[J]. PLoS One, 2014, 9(1): e86984. doi: 10.1371/journal.pone.0086984 [50] Burkhardt S, Zondervan I, Riebesell U. Effect of CO2 concentration on C: N: P ratio in marine phytoplankton: a species comparison[J]. Limnology and Oceanography, 1999, 44(3): 683−690. doi: 10.4319/lo.1999.44.3.0683 [51] Woodger F J, Badger M R, Price G D. Regulation of cyanobacterial CO2-concentrating mechanisms through transcriptional induction of high-affinity Ci-transport systems[J]. Canadian Journal of Botany, 2005, 83(7): 698−710. doi: 10.1139/b05-050 [52] Li Futian, Fan Jiale, Hu Lili, et al. Physiological and biochemical responses of Thalassiosira weissflogii (diatom) to seawater acidification and alkalization[J]. ICES Journal of Marine Science, 2019, 76(6): 1850. doi: 10.1093/icesjms/fsz028 [53] Taucher J, Jones J, James A, et al. Combined effects of CO2 and temperature on carbon uptake and partitioning by the marine diatoms Thalassiosira weissflogii and Dactyliosolen fragilissimus[J]. Limnology and Oceanography, 2015, 60(3): 901−919. doi: 10.1002/lno.10063 [54] Hancock A M, Davidson A T, McKinlay J, et al. Ocean acidification changes the structure of an Antarctic coastal protistan community[J]. Biogeosciences, 2018, 15(8): 2393−2410. doi: 10.5194/bg-15-2393-2018 [55] Wu Yaping, Campbell D A, 111Irwin A J, et al. 111Ocean acidification enhances the growth rate of larger diatom111s[J]. Limnology and111 Oceanography, 2014, 59(3): 1027−1034. doi: 10.4319/lo.2014.59.3.1027 [56] Li Futian, Wu Yaping, Hutchins D A, et al. Physiological responses of coastal and oceanic diatoms to diurnal fluctuations in seawater carbonate chemistry under two CO2 concentrations[J]. Biogeosciences, 2016, 13(22): 6247−6259. doi: 10.5194/bg-13-6247-2016 [57] Hutchins D A, Walworth N G, Webb E A, et al. Irreversibly increased nitrogen fixation in Trichodesmium experimentally adapted to elevated carbon dioxide[J]. Nature Communications, 2015, 6(1): 8155. doi: 10.1038/ncomms9155 [58] Tong Shanying, Gao Kunshan, Hutchins D A. Adaptive evolution in the coccolithophore Gephyrocapsa oceanica following 1, 000 generations of selection under elevated CO2[J]. Global Change Biology, 2018, 24(7): 3055−3064. doi: 10.1111/gcb.14065 [59] Li Futian, Beardall J, Collins S, et al. Decreased photosynthesis and growth with reduced respiration in the model diatom Phaeodactylum tricornutum grown under elevated CO2 over 1800 generations[J]. Global Change Biology, 2017, 23(1): 127−137. doi: 10.1111/gcb.13501 -
 下载:
下载:
 点击查看大图
点击查看大图
计量
- 文章访问数: 485
- HTML全文浏览量: 567
- PDF下载量: 345
- 被引次数: 0
 京公网安备 11010502036328号 京ICP备17033152号
京公网安备 11010502036328号 京ICP备17033152号